bruno51 ha scritto:
In questo caso, credo sia giusto parlare della varietà o forma "nivea", perchè forma popolazioni importanti. Quando invece, come da noi, si trova un qualche esemplare nel mezzo di una stazione normalmente colorata, credo sia più giusto parlare di soggetti "apocromici".
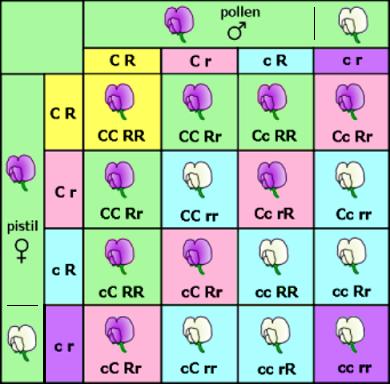
In senso strettamente biologico, è importante non confondere una pianta albina (completamente priva di clorofilla, foglie comprese) dalle forme albiflora (a fiori bianchi). Premesso questo, esiste in realtà una caratteristica comune tra le forme albiflora e l'albinismo vero e proprio: in entrambi i casi si tratta infatti di fenomeni geneticamente recessivi, ovvero un discendente potrebbe conservare i fiori bianchi, ma può anche sviluppare nuovamente fiori del colore standard. Charles Chamberlain Hurst (1870-1947) fu il primo a studiare le forme albiflora nelle orchidee. Egli scoprì che due geni sono responsabili per la colorazione dei fiori: il fattore C che inibisce la formazione del colore, e il fattore R che determina quale particolare colore verrà espresso (Rohrl, Helmut: For Heaven‘s Sake, It‘s Xanthic! Albinism in Orchids. In: Orchid Digest. Vol 69 (4), 2005. p.241). Ognuno di questi geni può esistere della forma inattiva, c e r. Le piante colorate eridatano uno o entrambi gli alleli attivi: CC e RR, Cc e RR, Cc e Rr o CC e Rr (forme a fiori rosa). Le forme a fiori bianchi hanno o cc o rr: CC e rr, Cc e rr, cc e RR, cc e Rr. C e R sono intesi come alleli dominanti che determinano la produzione di certi enzimi necessari per la produzione di pigmenti, tra cui le antocianine, che sono particolarmente importanti per il colore del fiore.
In alcune specie le forme a fiori bianchi sono abbastanza comuni e si possono sviluppare popolazioni stabili, in genere quando gli incroci avvengono ripetutamente solo tra piante a fiori bianchi. Si genera quindi una popolazione costituita interamente da piante con almeno un gene con entrambi gli allelli recessivi (cc o rr), andando a perdere gli individui eterozigoti (Cc o Rr) e omozigoti dominanti (RR o CC), che presenterebbero entrambi fiori rosa. Che sia presente un solo individuo o popolazioni intere non fa differenza sulla tassonomia, perchè in entrambi i casi (una o cento) si tratta sempre della stessa pianta, geneticamente parlando. Si tratta sostanzialmente di morfotipi che possono essere classificati al rango di forma. Quello che cambia non è la pianta in sè, quanto piuttosto i meccanismi che hanno portato alla selezione di quella determinata popolazione, che presenta solo quel particolare fenotipo, con perdita degli esemplari colorati. La cosa interessante e che a mio avviso andrebbe approfondita è proprio questa: quali sono i fattori che in quelle determinate aree hanno favorito la selezione di omozigoti recessivi (cc o rr) nei confronti degli omozigoti dominanti ed eterozigoti.
Ad oggi è stato dimostrato come in popolazioni di
Orchis mascula, la presenza di morfotipi bianchi aumenti la percentuale di impollinazione delle piante con colorazione normale dal 6% al 27%, e che alcune specie emettono differenti bouquet di profumi a secondo del morfotipo di colore:
Orchis mascula e
Anacamptis coriophora fragrans per esempio non hanno produzioni differenti di molecole volatili tra morfotipo bianco e colorato, mentre O. simia presenta differenti profili di molecole volatili tra i due morfotipi di colore. Se però si analizzano in modo approfondito i composti benzenoidi (chesono associati con laproduzione di antocianine) si vede che le infiorescenze bianche emettono più sostanze volatili ottenute da una particalare via metabolica (via dello shikimato o via dell'acido shikimico) rispetto alle piante colorate, sia in
O. mascula che in
O. simia. L'ipotesi corrente è che ci possano essere dei collegamenti tra colore e profumo floreale, influenzati probabilmente dagli stessi geni. Resta da stabilire in che modo la variazione intraspecifica di segnali floreali e colore influisca in realtà sull'attrazione degli insetti impollinatori e sul successo riproduttivo di queste orchidee.
Per concludere riporto alcuni taxa relativi a queste forme albiflora:
Anacamptis pyramidalis var.
albiflora Fors.-Major, Mém. Mus. Hist. Nat. 3: 33 (1817).
Anacamptis pyramidalis f.
albiflora Borg, Descr. Fl. Malt. Isl.: 72 (1927), nom. illeg.
Anacamptis pyramidalis lusus
albiflora (Fors.-Major) Soó, Bot. Arch. 23: 92 (1928).
Anacamptis pyramidalis var.
nivea P.Delforge, Naturalistes Belges 81: 396 (2000).
Anacamptis pyramidalis f.
nivea (P.Delforge) O.Gruss & M.Wolff, Orchid. Atlas: 34 (2007).
Bibliografia:L. Dormont, R. Delle-Vedove, et al., 2009: Rare white-flowered morphs increase the reproductive success of common purple morphs in a food-deceptive orchid. New Phytologist (2010) 185: 300–310. --->
PDF ONLINELaurent Dormont,1,* Roxane Delle-Vedove, 2010: Helping in food-deceptive orchids? A possible new mechanism maintaining polymorphism of floral signals. Plant Signaling & Behavior 5:5, 526-527. --->
PDF ONLINEB. Schatz, L. Dormont, R. Delle-Vedove: Presence, distribution and effect of white, pink and purple morphs on pollination in the orchid
Orchis mascula. European Journal of Environmental Sciences, Vol. 3, No. 2, pp. 119–128. --->
PDF ONLINEL Dormont, R Delle-Vedove, J-M Bessière, B Schatz, 2014: Floral scent emitted by white and coloured morphs in orchids. Phytochemistry; 100:51-9. --->
ABSTRACT ONLINE






